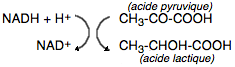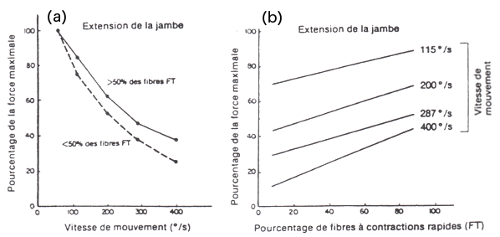MàJ
11/2013
Firefox, Safari
1440x900
La CIRCULATION SANGUINE
Généralités
La contraction cardiaque est assurée par les fibres du myocarde, réunies
en unités fonctionnelles. Le coeur est doué d'un automatisme lui
permettant de fonctionner spontanément suivant une fréquence cardiaque
propre. Cet automatisme, généré par le noeud sinusal, est sous la
dépendance des
systèmes parasympathique et
sympathique.
La capacité de polarisation et de dépolarisation du coeur est intrinsèque
(ne dépend pas du SNC), cependant le coeur est innervé par des
neuro-fibres qui peuvent en réguler le rythme (émotions...).
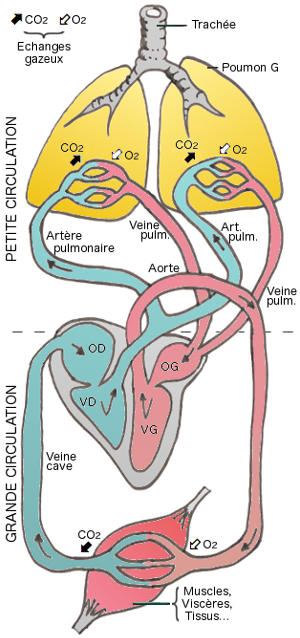
Le coeur est une pompe dont la fonction est d'éjecter le sang dans les
circulations pulmonaire et générale ou systémique (
petite
et grande circulations). Le retour veineux étant favorisé par les
effets concomitants de la fonction cardiaque, de la respiration, de la
gravité et de la contraction musculaire.
Le cycle cardiaque se décompose comme suit :
- une diastole auriculaire (ou remplissage auriculaire) à partir du
retour veineux des veines caves inférieure et supérieure pour
l'oreillette DR, de la circulation pulmonaire pour l'oreillette G
- une systole auriculaire (ou contraction des oreillettes) où le sang
est éjecté vers les ventricules à travers les valves
auriculo-ventriculaires qui se referment pour l'étape suivante
- une systole ventriculaire, qui éjecte le sang dans l'aorte
(ventricule G) vers le corps et le tronc pulmonaire (ventricule DR)
vers les poumons
Anatomie
Le coeur est situé dans le médiastin et repose sur la face sup. du
diaphragme. Mesurant environ 15 cm, il s'étend de la 2
ième côte
au 5
ième espace intercostale. Il est entouré et recouvert
partiellement des poumons.
Il n'est pas symétrique (2/3 de sa masse à gauche). Sa base supérieure,
regarde l'épaule droite et sa pointe (ou apex) est dirigée vers la hanche
gauche.
L'enveloppe du coeur est un sac à double paroi appelée péricarde :
- le péricarde fibreux lâche est composé de tissus conjonctifs denses.
Il protège le coeur et l'amarre au diaphragme et aux vaisseaux. Il
recouvre la 2ième paroi
- le péricarde séreux formé lui-même de 2 lames (lame pariétale
externe et lame viscérale plaquée contre le coeur) formant une cavité
remplie de liquide afin d'atténuer les frottements lors des battements
La paroi du coeur est formée de 3 tuniques, du DH vers le DD :
- l'épicarde
- le myocarde (muscle strié
capable de contraction)
- l'endocarde (cellules ramifiées et tissus conjonctif formant le
squelette fibreux) qui limite sa dilatation ainsi que la propagation
des influx nerveux
|
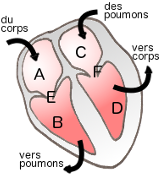
Il existe 4 cavités dans le coeur : 2 oreillettes dans sa partie
sup. et 2 ventricules dans sa partie inf. Deux cloisons les
séparent (septums inter-auriculaire et inter-ventriculaire)
- Les oreillettes :
lieu de petites contractions, elles ont une paroi mince et
sont de petites tailles. Elles participent peu à l'action de
pompage du coeur. Trois veines pénètrent dans l'oreillette
droite (A) : les veines caves sup. et inf. et le sinus
coronaire (sang provenant du myocarde). Les quatre veines
pulmonaires entrent dans l'oreillette gauche (C).
- Les ventricules :
Ils constituent la quasi totalité de la masse de coeur. Formés
des muscles papillaires, ils sont le point de départ de la
circulation. L'artère aorte démarre du ventricule gauche (D)
et donne naissance à la grande
circulation ; le tronc pulmonaire amène le sang aux
poumons en partant du ventricule droit (B) : c'est la petite
circulation. La paroi du ventricule G est 3 fois plus
épaisse que celle du ventricule DR.
- Valves tricuspide (E) et mitrale (F)
|
Dans le coeur, le sang circule en sens unique grâce au fonctionnement
de 4 valves anti-retour. La pression sanguine ouvrent et ferment ces
valves :
- 2 valves auriculo-ventriculaires (tricuspide à DR, mitrale à G)
empêchent le sang de refluer des ventricules vers les oreillettes
- 2 valves situées à la base de l'aorte et du tronc pulmonaires
empêchent le sang de remonter dans les ventricules lors de leur
décontraction
Le sang circule ensuite dans les vaisseaux :
- les artères quittent le coeur, puis se divisent en artérioles, ce
sont des zones de conduction
- les capillaires sont les zones d'échange (O2/CO2)
- les veinules puis les veines reconduisent le sang des capillaires
vers le coeur. Ce sang sera chargé de CO2 s'il provient de
la grande circulation ; d'O2
s'il provient de la petite circulation
Définitions
Petite
et grande circulations :
- Quand le sang circule entre le coeur et les poumons pour se
réoxygéner, il décrit une circulation pulmonaire ou petite
circulation.
- un fois oxygéné, il circule entre le coeur et les différents
organes conduisant l'oxygène et se chargeant en gaz carbonique
: c'est la circulation générale ou grande circulation
Sang
riche en O2 ; Sang
riche en CO2
Débit cardiaque (Qc) :
c'est la quantité de sang éjecté par le ventricule en fonction du
temps (1 minute). Les ventricules droit et gauche ont normalement
le même débit. Au repos, le débit cardiaque est d'environ 5 l/mn
et peut être multiplié par 5 à 6 par la pratique physique.
Qc=Fc x VES
Débit
cardiaque = fréquence cardiaque x volume d'éjection systolique
Fréquence
cardiaque (Fc) : l'augmentation de la fréquence
cardiaque augmente le débit cardiaque. Elle varie de 60 à 120
bat/mn, au-delà le temps de remplissage ventriculaire ainsi
que la perfusion coronaire diminuent. Valeur de la fréquence
cardiaque maximale théorique (FCmax en bat/mn) :
FCmax = 220-âge (Astrand et
Ryhming, 1954)
ou plus récemment : 191,5 - 0.007xâge² (Gellish, 2007)
La fréquence cardiaque augmente avec :
- L'hyperthermie
- L'activité physique
- L'anxiété
- L'hypotension
|
- L'hypoxémie
- L'anémie
- La douleur
- Les stimulants (tabac, café, médicaments...)
|
OD et OG : Oreillette droite et gauche
VD et VG : Ventricule droit
et gauche
|
|
Volume
d'éjection systolique (VES) : C'est la quantité de sang éjecté
par le ventricule lors d'une contraction. Au repos, le VES est de 60 à 100
ml par battement.
VES= Qc/Fc
Veines caves : Il existe 2 veines caves, l'une supérieure et
l'autre inférieure. Elles ramènent dans l'oreillette DR, le sang des
parties du corps respectivement situées au dessus et en dessous du
diaphragme.
Régulation du Rythme
A la base le rythme cardiaque est du à l'action indépendante d'un système
de commande intégré au coeur. Le système de conduction du coeur appeler
système cardio-necteur est composé de cellules cardio-nectrice (situées
dans le noeud sinusal). Leurs fonctions consistent à produire des influx
nerveux pour dépolariser le coeur des oreillettes aux ventricules. 70 fois
par minute, les cellules cardiaques vont se dépolariser créant un
potentiel d'action électrique qui se propage dans l'ensemble des cellules
musculaires cardiaques.

La longue période réfractaire du myocarde est de l'ordre de 250 ms
(milliseconde) empêche la sommation de secousses musculaires de type
tétanos.
Le coeur est prêt pour 3 milliards de battements (soit env. l'équivalent
d'une vie de 80 ans)
Le noeud sinusal, situé dans la paroi de l'oreillette DR, est le centre
rythmogène du coeur (rythme cardiaque ou sinusal), il se dépolarise
environ 70 fois par minute.
L'onde de dépolarisation (Cf.
animation) se
propage dans les oreillettes par les jonctions ouvertes puis arrive au
noeud auriculo-ventriculaire (situé au dessus de la valve
auriculo-ventriculaire dans la partie inférieur du septum
inter-auriculaire), l'influx y est retardé pendant 0,1s permettant aux
oreillettes de terminer leurs contractions avant que les ventricules ne
commencent la leur. La dépolarisation descend le long du faisceau de His
(ou auriculo-ventriculaire) qui se divise en 2 branches DR et G et
parcourent le septum inter-ventriculaire jusqu'à la pointe du coeur.
L'onde de dépolarisation termine son trajet dans les fibres de Purkinje
qui depuis l'apex remonte dans les parois des ventricules assurant leurs
contractions.
Le rythme cardiaque peut être modifié par le système nerveux autonome avec
les systèmes parasympathique (ralentisseur) et sympathique (accélérateur).
Rôle
des systèmes parasympathique et sympathique : Le système
parasympathique est le système extrinsèque prépondérant. Le noeud sinusal
a une activité automatique à 120 bat/mn qui est ralentit par le système
parasympathique à 70 bat/mn. Le système sympathique intervient lors d'un
effort physique afin d'adapter le débit cardiaque aux besoins de
l'organisme. Il joue aussi un rôle prépondérant en cas de stress (avec
décharge de catécholamines)
| Le
système parasympathique |
|
Le
système sympathique |
médiateur nerveux : nerf pneumogastrique (nerf X)
médiateur chimique : acétylcholine |
|
médiateur neurologique : le système sympathique
médiateur chimique : noradrénaline circulante
|
| Par
son action, il entraîne un effet : |
négatif
soit une bradycardie (diminution du rythme cardiaque) |
chronotrope
|
positif
soit une tachycardie (augmentation rythme cardiaque) |
négatif
soit une diminution de la contractilité cardiaque
|
inotrope
|
positif
soit une augmentation contractilité cardiaque |
négatif
soit une diminution de la conduction cardiaque
|
dromotrope
|
positif
soit une accentuation de la conduction cardiaque
|
négatif
soit une augmentation de l'excitabilité du myocarde |
bathmotrope
|
positif
soit une diminution de l'excitabilité du myocarde |
Electrocardiogramme
(ECG) : On peut étudier la variabilité de la fréquence cardiaque
(intervalle de temps qui sépare chaque battement) pour quantifier
l'activité des systèmes nerveux parasympathique et sympathique. L'ECG est
une représentation graphique de l'activité électrique du coeur. Son
tracé présente plusieurs "ondes". Les mesures habituellement effectuées
lors de l'analyse d'un ECG sont celles de l'onde P, de l'espace PR, du
complexe QRS et enfin de l'onde T :
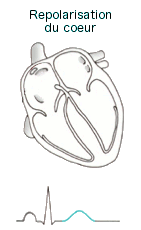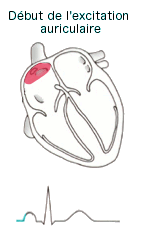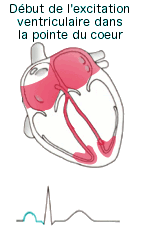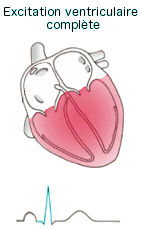 |
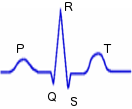 l'onde
P témoigne de la dépolarisation (et la contraction) des
oreillettes l'onde
P témoigne de la dépolarisation (et la contraction) des
oreillettes- l'espace PR correspond au temps de conduction de l'influx du
noeud sinusal au noeud auriculo-ventriculaire (conduction
auriculo-ventriculaire).
Sa durée normale, mesurée du début de l'onde P au début du
complexe QRS est de 0,12 à 0,20 seconde. La durée de l'espace
PR diminue lorsque la fréquence cardiaque augmente
- l'onde QRS ou complexe QRS (temps de conduction de l'influx
du noeud sinusal au noeud auriculo-ventriculaire) correspond à
la dépolarisation (et la contraction) des ventricules.
La forme et l'amplitude du QRS varient selon l'éventuelle
pathologie du muscle cardiaque
- l'onde T correspond à la repolarisation (la relaxation) du
coeur
|
En cas d'anomalie, le tracé doit idéalement être comparé avec un ECG plus
ancien du même patient, par ex. : une repolarisation ventriculaire
anormale n'a pas du tout la même signification si elle existe depuis
plusieurs années que si elle est récente.
Les Groupes Sanguins
| Groupe
sanguin |
Rhésus |
Répartition
mondiale |
Rang |
| 0 |
+ |
38% |
1° |
| - |
7% |
4° |
| A |
+ |
34% |
2° |
| - |
6% |
5° |
| B |
+ |
9% |
3° |
| - |
2% |
7° |
| AB |
+ |
3% |
6° |
| - |
1% |
8° |
|
La
répartiion mondiale montre de grandes fluctuations en fonction des
pays ou plutôt des continents, ainsi :
- en Europe on retrouve plus de 40% de O
pour autant de A
- Chez les indiens d'Amérique : 96% de O
(100% de O chez les
indien su Pérou)
- Certaines Tribus mongoles de Russie (Oyirads) : 41% de B,
26% de O, 23% de A
et 11% de AB
|
La RESPIRATION
Généralités
Physiologie de la respiration : La respiration (ou ventilation) est
l'ensemble des phénomènes qui concourent à assurer les échanges gazeux
entre le milieu ambiant et la cellule vivante. Plusieurs solutions ont été
adoptées de la cellule à l'être vivant : chez les mammifères, il s'agit
d'un système fermé dans lequel circule un "pseudo milieu extérieur".
L'appareil respiratoire a plusieurs rôles, voici les principaux :
-
Oxygénation tissulaire & élimination du gaz carbonique (Conduire
l'air chargé d'O2 du milieu extérieur au fond des alvéoles
et conduire l'air chargé de CO2 en sens inverse)
- Maintien du pH sanguin à une valeur moyenne
- Thermorégulation & balance hydrique
- Phonation
- (défense de l'organisme, fonction métabolique, filtre circulatoire,
réservoir sanguin)
Description
Les poumons forme l'interface entre le milieu externe et le sang, grâce à
leurs réseaux de capillaires au niveau desquels des échanges par diffusion
sont possibles.
Les voies aériennes supérieures conduisent l'air et comprennent :
- le nez (et la bouche), les fosses nasales, qui conditionnent l'air
en température jusqu'à à 37°C et en humidité, le sature en vapeur
d'eau ; mais aussi qui filtrent et purifient l'air à l'aide aux poils
et au tapis muco-ciliaire
- le pharynx (plus d'une vingtaine de muscles qui rigidifient les
voies aériennes à l'inspiration)
- le larynx (zone de rétrécissement, empêche les aliments de faire
"fausse route" et freine la sortie de l'air à l'expiration)
Enserré dans la la cage thoracique (
colonne
vertébrale,
sternum,
côtes),
les voies aériennes inférieures ont un rôle de conduction et de transition
:
- (la conduction) la trachée (12 cm de long. et 1,8 cm de diam.)
conduit et conditionne l'air en température, en humidité, qui se
divise pour formé l'arbre bronchique
- les bronches vers les poumons DR et G
- les ramifications bronchiques (bronchioles terminales et
respiratoires)
- (la transition) les sacs alvéolaires, alvéoles : se sont les unités
fonctionnelles pulmonaires où se font les échanges gazeux par
diffusion car leur paroi très mince est accolée à la paroi des
capillaires de la circulation pulmonaire
A l'expiration l'air fait le chemin inverse.
Lorsque
l'air se trouve dans les tuyaux, il ne sert à rien, une fois arrivé aux
alvéoles, seul l'oxygène sera envoyé dans le sang, les autres composé de
l'air seront dirigés en sens inverse par le mécanisme d'expiration afin
d'être expulsés.
Chez l'adulte, il existe 300 millions de sacs alvéolaires par poumons,
pour une superficie moyenne de 80 m
2 de surface d'échange
(cependant, il existe une grande variabilité interindividuelle). La
fréquence respiratoire moyenne au repos est de 14 respiration par minute.
La Plèvre
C'est une enveloppe fine composée de 2 feuillets qui recouvre les poumons
: le feuillet pariétal tapisse la paroi thoracique alors que le feuillet
viscérale colle à la paroi des poumons. Entre ces 2 feuillets, un liquide
dans la cavité pleurale, réduit la friction entre les poumons et la cage
thoracique lors de la respiration.
Inspiration
Elle correspond à l'ouverture thoracique, soit l'entrée de l'air dans les
poumons. Elle est essentiellement active sur le plan musculaire.
- étape "abdominale" : contraction des fibres musculaires du
diaphragme qui se traduit par un abaissement du centre phrénique
(aplatissement de la coupole), créant une dépression au niveau
thoracique et l'appel d'air dans les poumons. Ce mouvement de descente
du diaphragme (en réalité : aplatissement de la coupole)crée une
dépression au niveau du thorax qui appel l'air.
- étape "thoracique" : la descente du centre phrénique se stabilise,
les fibres musculaires du diaphragme poursuivant leur contraction,
engagent une élévation des côtes basses, ouvrant de la partie
inférieure du thorax en largeur. Ensuite les intercostaux externes et
élévateurs des côtes complètent l'ouverture thoracique avec
l'élévation des côtes supérieures.
Expiration
C'est la fermeture ("re-fermeture") de l'espace thoracique : l'air ressort
des poumons qui se vident. L'expiration peut se faire en 2 phases, selon
l'amplitude et selon le mode (passif ou actif).
- mode passif (normal) : les muscles qui s'étaient contractés à
l'inspiration se relâchent. Passivement, le thorax se referme avec un
abaissement des côtes, l'effet d'élasticité pulmonaire (retour de
l'« étirement ») et costale (détorsion) ainsi que l'effet de
la gravité. La relaxation du diaphragme lui redonne une forme plus
concave (remontée du centre phrénique)
- mode actif (respiration profonde) :
- dans un 1ier temps : contraction des expirateurs
costaux
- 2ième temps : contraction de la sangle abdominale
Il
existe une réelle complicité entre le diaphragme (au coeur de l'acte de la
respiration) et le transverse (centre musculaire de la posture).
La respiration représente le premier échange entre l'individu et son
milieu et concerne la globalité du corps. Il y a interdépendance entre
mobilité squelettique et mobilité respiratoire. En effet, de par la mise
en oeuvre de musculatures communes, les mobilités squelettique et
respiratoire sont intimement liées : tout mouvement corporel doit être
façonné en prenant sont origine dans celui de la respiration.
Muscles respirateurs
|
Muscles
inspiratoires |
 |
Muscles
expiratoires |
- Le diaphragme participe
à l'inspiration, en se contractant, il s'abaisse dans la
cavité abdominale
- Intercostaux externes : fibres obliques de bas en haut vers
l'ARR, soulèvent les côtes
Augmentation des diamètres vertical, antéro-postérieur et
latéral de la cage thoracique en même temps que les côtes
s'horizontalisent.
Inspiration amplifiée : muscle accessoire :
|
Diminution des diamètres vertical, antéro-postérieur et
cotelatéral de la cage thoracique en même temps que les côtes se
verticalisent.
Expiration forcée :
- Intercostaux internes : fibres obliques de bas en haut vers
l'AV abaissent les côtes
- serrage de la sangle abdominale (obliques, transverse),
carré des lombes
|
 Les
Volumes
Les
Volumes
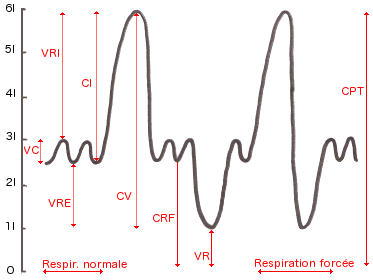
- Volume courant (VC) : volume qui entre ou qui sort des poumons à
chaque cycle respiratoire normal (environ 500ml)
- Volume de réserve inspiratoire (VRI) : volume qui est peut être
inspiré en plus de VC lors d'une inspiration forcée (2100 à 3200ml
selon taille du sujet)
- Volume de réserve expiratoire (VRE) : volume qui peut être expiré en
plus de VC lors d'une expiration forcée (1000 à 1200 ml)
- Volume résiduel (VR) : volume restant dans les poumons après une
expiration forcée (1000 à 1500ml). Ce volume résiduel joue un rôle
fondamental empêchant l'affaissement des poumons sur eux-même
Les Capacités
- Capacité inspiratoire (CI) : capacité totale d'air inspiré après une
expiration normale, CI = VC + VRI
- Capacité résiduelle fonctionnelle (CRF) : quantité d'air restant
dans les poumons après une expiration normale, CRF = VR + VRE (=2000 à
2500 ml)
- Capacité vitale (CV) : quantité d'air échangeable, VC + VRI + VRE (=
4000 à 4500 ml)
- Capacité pulmonaire d'air totale (CPT) : somme de tous les volumes
pulmonaires CV + VR (= 5000 à 6000 ml)
Les espaces Morts
Ce sont les espaces qui reçoivent de l'air mais qui ne participent pas aux
échanges avec le milieu intérieur. Espace mort total = espace mort
anatomique + espace mort alvéolaire :
- L'espace mort anatomique représente le volume de l'ensemble des
conduits du nez aux alvéoles
- L'espace mort alvéolaire représente les alvéoles qui ne participent
plus aux échanges (obstruées, affaissées ou non capillarisées)
Le METABOLISME ENERGETIQUE
Généralités
La dépense énergétique des muscles squelettiques peut varier de façon très
importante lors d'épreuves maximales : elles peuvent être multipliés par
80 à 100. Compte tenu de leur place importante dans le corps humain (1/3 à
2/5 de la masse totale), l'activité musculaire est directement liée à son
approvisionnement en énergie. L'énergie libérée par les aliments n'est pas
directement utilisée par l'organisme. Celle-ci est employée dans la
fabrication d'un autre composé chimique, l'Adénosine-Tri-Phosphate (ATP)
qui est emmagasinée dans toutes les cellules. C'est la dégradation de
l'ATP (débit d'énergie interne) qui fournit l'énergie utilisée pour la
contraction musculaire (débit d'énergie externe).
- Quels sont les mécanismes énergétiques qui fournissent de
l'énergie ?
- Comment la cellule musculaire est-elle réapprovisionnée en
ATP ?
Aspect Physiologique de la Production d'Energie
Le rôle central de l'ATP dans les échanges énergétiques des systèmes
biologiques a été perçu dès 1941 par Lipmann et Kalchar.
Plus qu'un descriptif approfondi des phénomènes bioénergétiques internes
et des relations biochimiques qui unissent les cellules fournissant de
l'énergie à celles qui en consomment; nous exposerons les différents
contextes des échanges énergétiques afin de caractériser :
- leur inertie (ou rapidité d'entrée en action, délai nécessaire afin
que le système entre en jeu d'une manière prépondérante pour assurer
l'essentiel de l'apport énergétique)
- leur débit maximal ou puissance (c'est à dire le débit d'énergie ou
quantité de molécules d'ATP par unité de temps). L'unité de puissance
est le Watt ou le Joule par seconde J/s (1 J/s =
1 Watt)
- leur capacité ou réserve totale d'énergie disponible (quantité
totale de molécules d'ATP produites). L'unité de mesure est le Joule
ou, généralement en physiologie, la calorie (1 cal =
4,186 Joules)
Au repos, les besoins énergétiques, au niveau du muscle strié
squelettique, sont assurés par l'hydrolyse de l'ATP en permanence
reconstituée grâce à l'énergie provenant de l'oxydation des nutriments.
ATP + H2-->O
Pin + ADP + Travail mécanique + Chaleur
Pin :
Phosphate inorganique, ADP : Adénosine-Di-Phosphate
Le "turnover" de l'ATP est important, c'est à dire que l'ATP, ADP et l'AMP
(Adénosine-Mono-Phosphate) sont inter-convertibles selon une réaction
catalysée par l'enzyme Myokinase (ou adénylatekinase) :
2 ADP <--> AMP + ATP
Au cours de l'effort, bien que considérablement augmentés (jusqu'à 100
fois), les besoins énergétiques sont toujours assurés par l'ATP. Le
système musculaire ajuste la production d'énergie en fonction des
modifications de la demande (Hultman et Sjöholm, 1983a). L'exercice
musculaire ne peut donc se produire que si l'ATP est continuellement
régénérée à partir de l'ADP.
Le muscle dispose de réserves d'énergie sous forme de phosphocréatine
(PCr), de glucides, de lipides et de substrats protidiques utilisés dans
des conditions particulières. La dégradation de ces substrats énergétiques
fournit l'énergie nécessaire aux processus contractiles.
| Substrats |
Quantité
(en kg) |
Energie
disponible
(en kJ) |
| Triglycérides |
12 |
446500 |
| Protéines (acides aminés
libres) |
6 |
78250 |
Glycogène :
hépatique
musculaire
|
0,060
0,400 |
638
4252 |
| Substrats circulants
(glucose, acide gras...) |
0,023 |
420 |
| PCr |
0,087 |
17 |
| ATP |
0,076 |
5 |
Répartition
des substrats énergétiques disponibles
chez un homme de 70 kg ayant 25 kg de muscles (d'après
Rieu, 1988)
Le délai d'intervention des processus de dégradation est variable, de
sorte que certains d'entre eux ne sont pas d'emblée disponibles. Au début
de l'exercice, c'est l'ajustement tardif de la consommation d'oxygène, dû
à l'inertie de l'adaptation cardio-respiratoire, qui a mis en évidence une
période transitoire pendant laquelle le muscle dépense plus que l'énergie
fournie par le métabolisme aérobie (Concept de dette d'oxygène introduit à
la suite des travaux de Hill, puis développé par Margaria). Ce mécanisme
énergétique est appelé anaérobie. Il est, lui même, subdivisé en deux
processus : l'anaérobie alactique et l'anaérobie lactique.
Métabolisme Anaérobie Alactique
On peut négliger l'apport énergétique lié au stock d'oxygène disponible
sous forme d'oxyhémyoglobine (tout au moins à basse altitude), ou
d'oxygène dissous et sous forme d'oxyhémoglobine dans les capillaires
musculaires.
En revanche, l'utilisation de l'ATP est immédiate. Ses faibles réserves
musculaires, environ 4,5 à 5,5 mMol.kg
-1 de muscle frais,
(Bangsbo et coll., 1990) (5 à 8 mMol.kg
-1 pour Saltin et
Gollnick, 1983) n'assurent que quelques contractions et la poursuite de
l'exercice exige son réapprovisionnement, au fur et à mesure de son
hydrolyse.
Un premier processus évite la chute de la concentration d'ATP. La
resynthèse de l'ATP s'effectue, dès les premières secondes de la
contraction, par le transfert d'une liaison phosphate d'un autre composé à
"haute énergie", la phosphocréatine (PCr), vers l'Adénosine-Di-Phosphate
(ADP) :
PCr --> Pin + Cr + énergie
Pin
: phosphate inorganique, Cr : créatine |
énergie + ADP + Pin --> ATP
enzyme :
créatine-phospho-kinase (CPK) |
Ces deux réactions sont couplées et se résument ainsi : ADP+PCr
--> ATP+Cr
La concentration de PCr est plus élevée dans les fibres à contractions
rapides (Essen et coll., 1975; Edstrom et coll., 1982) mais sa resynthèse
qui nécessite de l'oxygène est plus rapide dans les fibres musculaires à
contractions lentes, au contenu mitochondrial et à l'activité oxydative
plus importants (Saltin et Gollnick, 1983).
D'autre part, le mécanisme de l'ADP contribue aussi à la resynthèse
anaérobie de l'ATP :
2 ADP --> AMP+ATP
enzyme
: Myokinase
Mais l'apport énergétique de cette réaction reste encore mal connu. Elle
permettrait d'équilibrer les concentrations en AMP, ADP et ATP.
Spécificités du Processus Anaérobie Alactique
- Inertie : très faible et la puissance maximale est atteinte presque
instantanément
- Puissance : très élevée (jusqu'à 800 kJ.min-1 soit
13 kW), en particulier chez les sportifs qui possèdent une
prépondérance de fibres blanches rapides (IIa et IIb) riches en CPK
- Capacité : faible car la quantité de phosphocréatine est peu
importante, 15 à 20 mMol.kg-1 selon les auteurs et
n'autorise plus la resynthèse d'ATP pour des exercices réalisés à
puissance maximale, au-delà de quelques secondes (env. 5 s)
Ce métabolisme ne nécessite pas la présence d'oxygène et ne s'accompagne
pas de la formation d'acide lactique ; en revanche, la re-synthèse de PCr
nécessite de l'oxygène.
Métabolisme Anaérobie Lactique
Dès le début de l'exercice, la présence de catabolites ADP, AMP active les
enzymes du catabolisme du glycogène intramusculaire (Glycogénolyse) et du
glucose libre cellulaire ou sanguin (Glycolyse). Ces voies métaboliques
conduisent à la production de lactate.
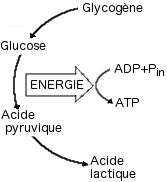 |
Le
système de
l'acide lactique
(d'après Fox et
Mathews, 1984)
|
LA GLYCOGENOLYSE : C'est la
dégradation du glycogène en glucose-1-phosphate (G-1-P) :
Glycogène <->
Glycogène + G-1-P
(n
glucose)
(n-1 glucose)
Cette réaction est catalysée par une enzyme complexe, la glycogène
phosphorylase, elle même secondée par la glycogène synthétase. Ces deux
enzymes existent sous deux formes interconvertibles, leur régulation est
complexe et dépend de nombreux facteurs (Gross et Mayer, 1974; Gollnick et
coll., 1978; Richter et coll., 1982).
LA
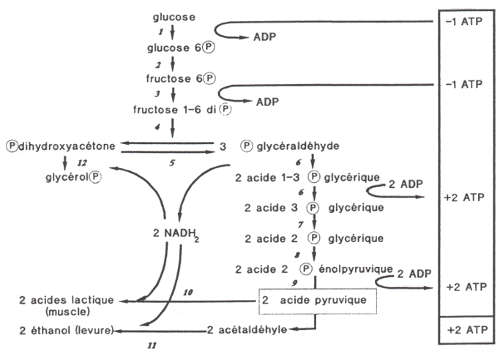
GLYCOLYSE : C'est la voie métabolique de production d'ATP qui
résulte de la dégradation du glucose. En absence d'oxygène, le catabolisme
du glucose en acide pyruvique est effectué par l'action séquentielle de 10
enzymes assurant la synthèse de 4 molécules d'ATP mais en consommant 2,
soit un solde de 2 molécules d'ATP.
|
Les
étapes de la glycolyse
(d'après Kruh, 1989)
A partir de l'étape 6,
un glucide à 6 carbones
est scindé en 2 glucides
3 carbones, de sorte que
le nombre de molécules à
chaque étape ultérieure
est doublé. Les étapes 6
sont responsables de la
synthèse nette de molécules
d'ATP et de NADH qui
nécessite un apport en NAD |
L'enzyme phospho-fructo-kinase (PFK) a été identifiée comme le régulateur
de cette voie métabolique (Newsholme et Start, 1974). L'enchaînement de
ces nombreuses réactions s'accompagne de la libération d'hydrogène et
nécessite la présence d'un accepteur d'hydrogène, la
nicotinamide-adénine-dinucléotide (NAD
+) qui est réduite en
NADH + H
+ :
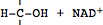
<-->
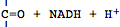 Réduction
du NAD (librement adapté de Alberts et coll., 1988)
Réduction
du NAD (librement adapté de Alberts et coll., 1988)
La poursuite de la glycolyse anaérobie exige une concentration stable de
NAD. Le NADH
2 doit céder son hydrogène pour être de nouveau
disponible. Si la cellule dispose d'oxygène, l'oxydation de la NADH
2
s'accomplit au niveau du cycle de Krebs dans les mitochondries. Si
l'apport en oxygène est insuffisant, la régénération du NAD
+
s'effectue, par une réduction de l'acide pyruvique en acide
lactique :
D'après les travaux de Sahlin et coll. (1987) le rapport entre le lactate
et le pyruvate serait proportionnel au rapport entre le NADH
2
et le NAD
+ (équilibre de l'oxido-réduction). Le flux de
formation de l'acide lactique serait associé, durant l'exercice physique,
à la réoxydation cytoplasmique du NADH
2 (Katz et Sahlin, 1988).
En définitive, la synthèse de l'ATP s'accompagne de la production de deux
molécules de lactate par molécule de glycogène dégradé. Cette voie
métabolique aboutit à la formation, puis à l'accumulation lactique
progressive dans la cellule qui participe à l'acidose musculaire.
L'acidose graduelle peut amener le pH à 6,5 ce qui altère les
propriétés contractiles des cellules musculaires et inhibe l'action des
enzymes de la glycolyse.
Spécificités du Processus Anaérobie Lactique
- Inertie : pas négligeable. Plus d'une dizaine d'enzymes sont
nécessaires à la formation d'acide lactique et certaines doivent être
activées (comme la PFK). Les nombreuses transformations intermédiaires
aboutissent à un délai d'intervention efficace qui se situait entre
vingt et trente secondes pour Thill et coll. (1982) mais qui est
aujourd'hui mesuré dès la 6ième seconde d'un sprint sur
cyclo-ergomètre (Mercier et coll., 1991)
- Puissance : dépend des enzymes de la glycolyse. Moins importante
que celle du métabolisme anaérobie alactique, elle varie de 200 à
250 kJ.min-1 chez le sujet non entraîné (soit environ
3,5 kW), et peut atteindre 500 kJ.min-1 chez le
sportif spécialiste de courses dont la durée varie de 1 à 2 minutes
- Capacité de la glycolyse anaérobie : grossièrement proportionnelle
à la concentration de lactate tolérée par l'organisme, elle même
proportionnelle à la concentration sanguine en lactate. Elle
représente 14 à 17 mMol.l-1 chez le sédentaire (soit
environ de 75 kJ), prés du double chez certains athlètes. Le
nombre de molécules d'ATP resynthétisées par ce processus n'autorise
des exercices intenses que pendant une durée d'environ 40 secondes
Les activités sollicitant ce métabolisme sont soient, des activités
intenses soutenues plusieurs dizaines de secondes; soient, des activités
très intenses de courte durée, mais répétées avec un intervalle de
récupération insuffisant. Dans ce dernier cas, les réserves de
phosphocréatine s'épuisent au fur et à mesure des répétitions et la
glycolyse anaérobie est activée.
Quand
l'intensité d'effort augmente, les muscles produisent de plus en plus
d'acide lactique, lequel est rapidement dissocié en proton (H
+)
et en anion lactate (C
3H
5O
3-).
La baisse du pH, si elle est important, peut altérer la contraction
musculaire. En revanche, une fois libéré, l'anion lactate peut être oxydé
in-situ par les muscles ou par d'autres tissus/cellules (neurones,
coeur...) notamment par l'intermédiaire de transporteurs de
monocarboxylate. Une mole de lactate oxydé dans la mitochondrie permet de
générer 17 moles d'ATP. Pendant la récupération, l'anion lactate permet de
re-générer les stocks de glycogène dans le foie (Cycle de Cori).
Métabolisme Aérobie
Le métabolisme aérobie n'est que succinctement traité dans ce chapitre.
C'est donc l'apport en oxygène, qui rend possible les efforts de longue
durée, grâce à l'énergie fournie par l'oxydation principalement des
glucides et des lipides, au niveau de la mitochondrie.
Les différentes réactions de ce métabolisme sont généralement
différenciées en 3 étapes : la glycolyse aérobie (qui présente un
parallèle certain avec la
glycolyse anaérobie),
le cycle de Krebs et le système de transport des électrons (Fox et
Mathews, 1984).
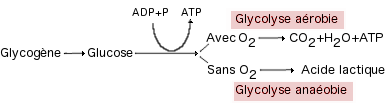 |
La
glycolyse anaérobie et aérobie
Sans oxygène, l'acide
pyruvique
se transforme en acide lactique
sans autre resynthèse d'ATP.
Avec oxygène, l'acide pyruvique
entre dans le cycle de Krebs |
L'aérobie est à la base des exercices de longue durée pour trois
raisons :
- la réserve d'énergie est importante
- le rendement énergétique de l'oxydation des lipides et des glucides
est élevé
- il n'y a pas d'accumulation de catabolites qui pourrait perturber le
métabolisme, en effet, le gaz carbonique est éliminé par les poumons,
et l'eau par la sueur et la respiration
Le bilan énergétique de l'oxydation des glucides est nettement plus
favorable que celui de la glycolyse anaérobie. Chaque molécule de glucose
produit 38 molécules d'ATP selon une succession de réactions couplées que
l'on résume ainsi :
C6H12O6 + 6 O2
--> 6 CO2 + 6 H2O + Energie
C6H12O6
: unité glycosidique
Energie + 38 ADP + 38 Pin
--> 38 ATP
Pin :
Phosphate inorganique
L'oxydation des lipides épargne le glycogène musculaire et hépatique, de
plus les réserves en acides gras libres et triglycérides sont très
importantes (env. 12 kg chez un homme de 70 kg ayant 25 kg
de muscles). Le rendement de l'oxydation des lipides est élevé : 128
molécules d'ATP pour une molécule de lipide. Chez l'athlète entraîné pour
des activités sportives d'endurance, l'utilisation des acides gras libres
peut représenter jusqu'à 70 % de la dépense énergétique des exercices
de longue durée.
Enfin, une oxydation des substrats protidiques, acides aminés, peut avoir
lieu, mais elle n'entre en jeu que lors de dénutrition lipidique
importante ou après un effort largement supérieur à une heure. Elle ne
représente que 3 à 15 % de la dépense totale d'énergie. Son rendement
est proche de celui de la glycogénolyse.
Spécificité du Processus Aérobie
- Inertie : très importante, sous la dépendance de l'activation de la
chaîne respiratoire mitochondriale, de l'adaptation des fréquences et
débits cardiaques et ventilatoires (plusieurs dizaines de secondes à
quelques minutes sont souvent nécessaires pour atteindre un équilibre
entre besoins et apports en oxygène), ainsi que les nombreuses
réactions qui assurent le catabolisme des substrats utilisés
- Puissance : inférieure à anaérobie lactique ou alactique. La
puissance maximale résulte de la quantité maximale d'oxygène qui peut
être utilisée par les mitochondries, elle est définie par la
consommation maximale d'oxygène. Les valeurs de référence couramment
utilisées sont de 40 ml.min-1.kg-1 pour un
homme de 70 kg (soit 60 kJ.min-1) et peuvent
doubler chez l'athlète de haut niveau
- Capacité : peut sembler illimitée car les réserves des substrats
utilisés sont importantes et l'oxygène peut être puisé dans
l'environnement. Il faut cependant relativiser cette notion en tenant
compte des 3 facteurs suivants :
- intensité du travail
- niveau d'entraînement du sujet
- capacité de thermolyse de celui-ci.
Synthèse
La contraction musculaire est donc étroitement liée à la décomposition de
l'ATP, elle-même resynthétisée différemment en fonction de la durée et de
l'intensité de l'exercice musculaire. Les processus qui aboutissent à la
formation d'ATP peuvent être comparées en ce qui concerne leurs
caractéristiques générales (substrats synthétisés, catabolites rejetés,
vitesses de mise en jeu et l'utilisation ou non d'O
2) et
peuvent être résumés ainsi :
La connaissance des réponses énergétiques de l'organisme, en fonction du
type de tâche à accomplir, autorise l'approfondissement des compétences en
matière d'adaptation de l'entraînement. Le tableau suivant précise le
délai d'intervention, la puissance et la capacité des différents processus
métaboliques. Il prend en compte des valeurs les plus souvent citées dans
la littérature.
|
Anaérobie
alactique
|
Anaérobie
lactique
|
Aérobie
|
| Substrats utilisés |
ATP+ PCr |
glycogène et glucose |
lipides, glucides
(et protides)
|
| Délai
d'intervention |
nul |
6 à 30 s
|
1'30" à 3
min |
Puissance
ou débit d'Energie |
400 à 750 kJ/min* |
200 à 500 kJ/min* |
60 à 120 kJ/min* |
Capacité
ou quantité d'Energie |
très
faible
30 à 50 kJ* |
faible
95 à 120 kJ* |
très
élevés
dépend de VO2max* |
* dépend
des caractéristiques individuelles et du niveau de pratique sportive
(librement adapté de Thill et coll., 1982; Mercier et coll., 1991)
Longtemps les
3 systèmes énergétiques
ont été décrits comme s'enchaînant dans le temps. Il existe cependant une
interaction entre les sources aérobie et anaérobies au cours de
l'exercice. De même, la séparation entre périodes alactique et lactique ne
doit pas être aussi tranchée qu'on le supposait. Jacobs et coll. (1983a),
Jones et coll. (1985), Mercier et coll. (1989, 1991) montrent une hausse
significative de la concentration des lactates musculaires ou sanguins
après un exercice intense de 6 à 10 secondes. D'autre part, Hultman et
Sjöholm (1983b) ont calculé une participation de 20% de la glycolyse, dès
la première phase d'une contraction maximale induite par stimulation
électrique.
Bibliographie
ALBERTS B., BRAY D., LEWIS J., RAFF M., ROBERTS K., WATSON
J.D. (1988) Biologie moléculaire de la cellule. Traduit de
l'américain par M. MINKOWSKI. 5° ed. Flammarion (Paris), 67-75.
BANGSBO J., GOLLNICK P.D., GRAHAM T.E., JUEL C., KIENS B., MIZUNO M.,
SALTIN B. (1990) Anaerobic energy production during
exhaustive exercise in humans. J. Physiol. (London) 422,
539-559. EDSTROM L., HULTMAN E., SAHLIN K., SJÖHOLM H.
(1982) The contents of high energy phosphate in different fiber types in
skeletal muscles from rat, guinea-pig and man. J. Physiol. Lond.
332, 47-58. ESSEN B., JANSSON E., HENRIKSSON J., TAYLOR A.W.,
SALTIN B. (1975) Metabolic characteristics of fiber types in
human skeletal muscle. Acta Physiol. Scand. 95, 153-165. FOX
E.L., MATHEWS D.K. (1984) Bases physiologiques de l'activité
physique. (Traduit et adapté par F. PERONNET de Physiological basis of
physical education and athletics, 1981, WB. Saunders Compagny,
Philadelphia). Decarie ed. Montréal. ed. Vigot, (Paris). pp 404. GOLLNICK
P.D., KARLSSON J., PIEHL K., SALTIN B. (1978) Phosphorylase
in a human skeletal muscle during exercise and electrical stimulation. J.
Appl. Physiol. Respirat. Enviro. Exercise Physiol. 45, 852-857. GROSS
S.R., MAYER S.E. (1974) Regulation of phosphorylase b to a
conversion in muscle. Life Sci. 14, 401-414. HULTMAN
E., SJÖHOLM H. (1983a) Energy métabolism and contraction
force of human skeletal muscle in situ during electrical stimulation. J.
Physiol. Lond. 345, 525-532. HULTMAN E., SJÖHOLM H.
(1983b) Substrate availability. In : Biochemisrty of exercise.
H.G. KNUTTGEN, J. VOGEL, J. POORTMANS (Eds). Internat Series on
Sport Sciences. 13, 63-75. JACOBS I., BAR-OR O., DOTAN
R., KARLSSON J., TESCH P. (1983a) Changes in muscle ATP, CP,
Glycogen and lactate after of the Wingate test. In : Biochemistry
of Exercise. H.G. KNUTTGEN, J.A. VOGEL and J. POORTMANS (Eds), International
Series of Sport Sciences, Human Kinetics Inc. 13, 234-238.
JONES N.L., Mac CARTNEY N., GRAHAM T., SPRIET L., KOWALCHUCK
J.M., HEIGENHAUSER G.J.F., SUTTON J.R. (1985) Muscle
performance and metabolism in maximal isokinetic cycling at slow and
fast speeds. J. Appl. Physiol. 59, 132-136. KATZ A.,
SAHLIN K. (1988) Regulation of lactic acid production during
exercise. J. Appl. Physiol. 65, 509-518. KRUH J. (1989)
Production d'énergie. In : Biochimie, vol II Métabolismes.
Hermann ed. (Paris). MERCIER J., MERCIER B., PREFAUT C. (1989)
Sollicitation du métabolisme anaérobie lactique lors de la répétition
d'exercices brefs et intenses. C.R. Soc. Biol. 183, 60-66. MERCIER
J., MERCIER B., PREFAUT C. (1991) Blood lactate increase
during the force velocity execise test. Int. J. Sports Med.
12, 17-20. NEWSHOLME E.A., START C. (1974) Regulation
in metabolism. New-York, Wiley Interscience. RICHTER E.A.,
RUDERMAN N.B., GAVRAS H., BELUR E.R., GALBO H. (1982) Muscle
glycogenolyse during exercise : dual control by epinephrine and
contractions. Am. J. Physiol. E242 : E25-E32. RIEU
M. (1988) Bioénergétique de l'exercice musculaire et de
l'entrainement physique. P. CERRETELLI, C.Y. GUEZENNEC, K. HAINAUT, H.
HOWALD, P. PESQUIES, J. POORTMANS, P.E. Di PRAMPERO, D. WASSERMANN
sous la direction de M. RIEU. ed Puf (le biologiste). SAHLIN K,
KATZ A., HENRIKSSON J. (1987) Redox state and lactate
accumulation in human skeletal muscle during dynamic exercise. Biochem.
J. 245, 551-556. Saltin B., GOLLNICK
P.D. (1983) Skeletal muscle adaptability : Significance
for metabolism and perfformance. In : Handbook
of physiology. Section 10. Skeletal muscle 555-631
Baltimore : Williams & Wilkins. THILL E., THOMAS R.,
CAJA J. (1982) Manuel de l'éducateur sportif. Ed. Vigot
(Paris)
Introduction a la PHYSIOLOGIE
MUSCULAIRE
Généralités
L'évaluation des aptitudes physiologiques d'un individu (sportif ou
sédentaire) se caractérise principalement par l'étude de la capacité et de
la puissance énergétique externe qu'il peut développer lors d'un exercice
dont l'intensité et la durée font théoriquement appel au processus
énergétique interne à évaluer.
Nous ne parlerons pas ici en détail de la structure du muscle mais plutôt
de son comportement mécanique que nous envisagerons en terme de signal de
sortie musculaire.
Le muscle squelettique est composé de cellules allongées polynuclées, les
fibres musculaires. La fibre musculaire contient des
éléments
contractiles sous la forme de myofilaments composés de molécules
d'actine et de myosine ainsi que des protéines auxquelles elles sont
associées. La contraction musculaire est réalisée grâce au glissement des
protéines le long des filaments. Ce processus, qui provoque la mise en
tension et les variations de longueur, est un grand consommateur d'énergie
et dépend de l'
apport en ATP comme nous l'avons
explicité précédemment.
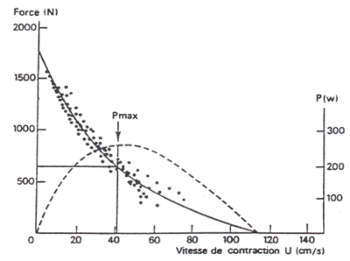
Relation Force-Vitesse
Premièrement la force développée par un muscle relève du nombre d'unités
motrices recrutées. D'autre part, la vitesse maximale à laquelle un muscle
se contracte dépend de la force qui lui est opposée. La vitesse maximale
absolue d'un mouvement ne peut être obtenue que pour des charges nulles.
Cette vitesse maximale est d'autant plus grande que les segments corporels
et les muscles qui les commandent sont petits.
Ce thème a fait l'objet de nombreux travaux depuis les premières
expériences de Fenn et Marsh (1935) et de Hill (1938) sur le muscle isolé,
liant sa vitesse de raccourcissement et sa force de contraction. Le
concept de proportionnalité, entre la force produite et la vitesse de
raccourcissement des fibres musculaires, a donné les bases du diagramme
force-vitesse selon une relation mathématique diversement décrite.
Diagramme
force-vitesse instantanée.
Contractions maximales du biceps brachial
(d'après Pertuzon et Bouisset, 1971) |
 |
Pour Fenn et Marsh, la relation entre ces deux grandeurs est
exponentielle, alors que Hill la décrit comme hyperbolique :
|
P = Po e-V/b - F
Fenn et Marsh, 1935
|
(P + a) V = b (Po
- P)
Hill, 1938 |
Po
est la tension isométrique maximale`
b est une constante ayant la dimension d'une vitesse
F est une force de friction faible de l'ordre de 3 à 5% de Po |
P
est la charge ; Po la
tension isométrique maximale
V est la vitesse de
raccourcissement
a et b sont des constantes (Close, 1972) ayant respectivement
les dimensions d'une force et d'une vitesse (Auber, 1969)
|
Ces deux équations montrent que
la force de contraction développée par un muscle diminue lorsque la
vitesse à laquelle il se raccourcit augmente. Il faut cependant
remarquer que ces modèles ne sont pas extrapolables pour des
allongements musculaires. La relation force-vitesse présente trois
caractéristiques à souligner :
- La force maximale développée par un muscle décroît à mesure que la
vitesse du mouvement augmente, elle ne s'observe donc que pour une
vitesse nulle
- La puissance mécanique de sortie que le muscle actif est capable de
délivrer est fonction de l'enveloppe de la courbe : P=FV
- La relation "structure/fonction" : Quelle que soit la force
développée, plus le muscle contient de fibres à contractions rapides (Fast
twitch), plus la vitesse est élevée.
Inversement, quelle que soit la vitesse du mouvement, la force de
contraction est d'autant plus élevée que le muscle contient un fort
pourcentage de fibres rapides (Cf. schéma ci-dessous ; d'après
les données de Coyle et coll., 1979)
(a)
Relation force-vitesse en fonction de la vitesse de mouvement et du
pourcentage de fibres à contractions rapides (FT).
(b) Quelle que soit la vitesse du mouvement, plus le pourcentage de
FT est élevé, plus la force développée est grande
Chez l'homme, la relation force-vitesse pour des mouvements
mono-articulaires simples, est, elle aussi, hyperbolique (Wilkie, 1950 ;
Komi, 1973) ou exponentielle (Mc Cartney et coll., 1985) suivant les
estimations mathématiques.
Il n'en est, cependant, pas de même
pour les mouvements complexes, polyarticulaires, impliqués lors
d'exercices mettant en jeu plusieurs groupes musculaires et lors
de mouvements polycycliques.
Dans ce cas, pour certaines conditions de vitesse, il existe une
relation linéaire
(Pirnay et Crielaard, 1979; Nadeau et coll., 1983) entre la
fréquence de pédalage et la force de freinage.
Cette linéarité peut être admise, sur cyclo-ergomètre mécanique,
pour des fréquences de 100 à 200 Tours par minute (Peres et coll.,
1981; Vandewalle et coll., 1983, 1988). |
|
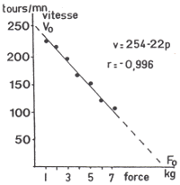 |
Vitesse
maximale de pédalage en fonction de diverses forces de freinage
(adapté de Vandewalle et coll., 1983) |
Puissance Mécanique d'un Groupe Musculaire
Lorsqu'une épreuve est susceptible de fournir les valeurs de la force mise
en jeu par un groupe musculaire, ainsi que la vitesse d'exécution de ce
mouvement, la puissance mécanique est obtenue par le produit de la force
par la vitesse.
Le pic de la courbe de
puissance représente une puissance mécanique maximale développée par le
groupe musculaire. Cette puissance maximale (Pmax), ou pic de puissance,
est généralement associée à une puissance énergétique maximale. Elle n'est
pas obtenue pour des valeurs maximales de force ou de vitesse, mais pour
le meilleur produit entre ces deux paramètres.
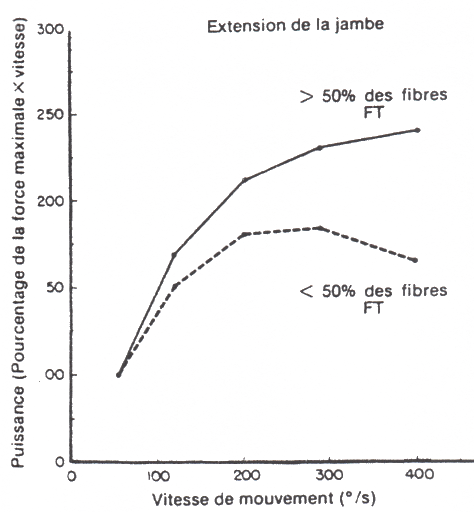
La relation puissance-vitesse est illustrée ci-dessous. Quelle que soit la
vitesse du mouvement, plus le pourcentage de fibres musculaires à
contractions rapides est élevé, plus la puissance maximale est importante.
Relations
entre la puissance et la vitesse de
mouvement en fonction du pourcentage de
fibres à contractions
rapides (FT) (d'après les données de
Coyle et coll., 1979)
En effet, les propriétés contractiles des fibres musculaires sont
différentes en fonction de leurs types de fibres. La classification de
Brooke et Kaiser, 1970 semble aujourd'hui la plus suivie :
- fibres lentes ou slow twitch : type I
- fibres rapides ou fast twitch : IIa et IIb
- fibres transitoires : IIc et IIab
La force relative est similaire, mais la vitesse de raccourcissement est 4
fois plus élevée pour les fibres rapides. La puissance de sortie
développée est respectivement estimée à 40 et 155 W.kg
-1
de muscle frais (Faulkner et coll., 1986). Les plus hautes valeurs de
puissance sont observées chez les sprinters qui possèdent un grand
pourcentage de FT, les plus faibles sont observées chez les marathoniens
au pourcentage de fibres lentes élevé.
Relation Energie Chimique / Energie Mécanique
L'énergie peut se convertir d'une forme à une autre (chimique, mécanique,
thermique...). L'exercice musculaire nécessite la conversion de l'énergie
chimique des aliments et des substrats énergétiques en énergie mécanique.
Nous pouvons associer certains types d'exercice musculaire à une filière
énergétique plus ou moins différenciée. Ainsi, pour un test qui mesure la
puissance anaérobie, on choisira des épreuves dont la durée est inférieure
à la minute, inférieure à quelques secondes pour des épreuves de type
essentiellement anaérobie alactique. Pour les tests de puissance aérobie,
des épreuves d'au moins 10 minutes sont généralement employées. La
puissance aérobie maximale est associée à la puissance mécanique
développée au moment de l'atteinte de la consommation maximale d'O
2.
De la même manière, les physiologistes de l'exercice physique, qualifient
de puissance anaérobie maximale, ou associent à celle-ci, la puissance
mécanique maximale mesurée lors du type d'épreuve mettant en jeu le
métabolisme anaérobie (Komi et coll., 1977a; Pirnay et Crielaard 1979;
Davies et Young, 1984; Inbar et Bar-or, 1986; Ferretti et coll., 1987;
Vandewalle et coll., 1987a, 1988; Mercier B. et coll., 1992).
En réalité, l'ensemble des tests du métabolisme anaérobie ne donne que des
indications fondées sur la connaissance de la rapidité de l'utilisation et
de l'épuisement des réserves de phosphagène. En effet, nous pouvons
penser, que ce sont les réserves d'ATP et de PCr, disponibles d'emblée,
qui fournissent l'énergie utilisée par les contractions musculaires
intenses de courte durée, mais aucune mesure directe n'en a apporté la
"preuve formelle" (Cazorla et coll., 1984).
Bibliographie
AUBER X. (1969) Le muscle strié, Kayser éd.,
Physiologie, Système nerveux et muscles, Flammarion, 1261-1323.
BROOKE MH., KAISER KK. (1970) Muscle fiber types: how many
and what kind? Arch. Neurol. 23, 369-379. CAZORLA
G., LEGER L., MARINI J.F. (1984) Les épreuves d'effort en
physiologie : épreuves et mesures du potentiel anaérobie. Travaux
et Recherche en EPS. 7, 82-94. CLOSE R.I.
(1972) Dynamic properties of mammalian skeletal muscle. Physiol.
Rev. 52, 129-191. COYLE E.F., COSTILL D.L., LEMES G.R.
(1979) Leg extension power and muscle fiber composition. Med.
Sci. Sports. 11, 12-15. DAVIES C.M.T., YOUNG K.
(1984) Effect of external loading on short term power output in
children and young male adults. Eur. J. Appl. Physiol. 52,
351-354. FAULKNER J.A., CLAFLIN D.R., Mac CULLY K.K.
(1986) Power output of fast and slow fibers from human skeletal
muscles. In : Human muscle power (eds) N.L. JONES, N.
Mac CARTNEY and A.J. Mac COMAS. Human Kinetics, Champaign
IL, 81-91. FENN W.O., MARSH B.S. (1935) Muscular
force at differente speeds of shortening. J. Physiol. Lond.
85, 277-297. FERRETTI G., GUSSONI M., Di PRAMPERO P.,
CERRETELLI P. (1987) Effects of exercices on maximal
instantaneous muscular power of humans. J. Appl. Physiol.
Vol.62 n°6, 2288-2294. HILL A.V. (1938) The heat of
shortening and the dynamic constant of muscle. Pro. R. Soc.
(London) 126, 136-195. INBAR O., BAR-OR O. (1986)
Anaerobic characteristics in male children and adolescents. Med.
Sci. Sports and Exerc. 18, 264-269. Komi P.V.
(1973) Measurement of force velocity relationship in human muscle
under concentric and excentric contractions. Med
and sport, vol. 8 : Biomechanics III Karger, Basel,
224-229. KOMI P.V., RUSKO H., VOS J., VINKO V.
(1977a) Anaerobic performance capacities in athletes. Acta
physiol.Scand. 100, 107-144. Mc CARTNEY N.,
OBMINSKI G., JONES N.L. (1985) Torque velocity relationship
in isokinetic cycling exercise. J Appl Physiol. 58,
1459-1462. MERCIER B., MERCIER J., GRANIER P., LE GALLAIS D.,
PREFAUT C. (1992) Maximal anaerobic power :
relationship to anthropometric characteristics during growth. Int.
J. Sports Med. 13, 21-26. NADEAU M.,
CUERRIER J.P., BRASSARD A. (1983) The bicycle ergometer for
muscle power testing. Can. J. Appl. Sports Sci. 8, 41-46. PERES
G., VANDEWALLE H., MONOD H. (1981) Aspect particulier de la
relation charge-vitesse lors du pédalage sur cyclo-ergomètre. J.
Physiol. (Paris) 77, 10A. PERTUZON
E., BOUISSET S. (1971) Maximum velocity of
movement and maximum velocity of muscle short-ening. Med
and sport, vol 1 : Biomechanics II, Karger (ed).
PIRNAY F., CRIELAARD J.M. (1979) Mesure de la
puissance maximale anaérobie alactique. Méd. Sport. 53,
13-16. VANDEWALLE H., PERES G., MONOD H. (1983)
Relation force-vitesse lors d'exercices cycliques réalisés avec les
membres supérieurs. Motricité Humaine. 2, 22-25. VANDEWALLE
H., HELLER J., PERES G., RAVENEAU S., MONOD H. (1987a)
Etude comparative entre le Wingate test et un force-vitesse sur
ergocycle. Sci. Sports. 2, 279-284. VANDEWALLE H.,
PERES G., HELLER J., MONOD H. (1988) Intérêts et limites
des relations forces-vitesses chez l'homme. Sciences et Motricité,
4, 38-46. WILKIE D.R. (1950) The relation between
force and velocity in human muscle. J. Physiol. (London).
110, 249-280